Rust Family Foundation: Archaeology Grants Program
Unveiling Ancient Maya Migration Histories:
Evidence from Stable Isotopes and aDNA
[RFF-2018-51]Principal Investigator:
Angelina J. Locker, The University of Texas at Austin
How ancestral veneration, social memory, migration, and genetics intersect is poorly understood in archaeological contexts. This research draws upon theories of social memory (Assmann and Czaplicka 1995; Chesson 2001; Connerton 1989; Gillespie 2010; Halbwachs 1980) to examine burial practices, migration, and kinship of the ancient Maya from the La Milpa Polity (LMP) of NW Belize via the study of body positioning, isotopes, and ancient DNA (aDNA). This research focuses on ancient Maya burials fro
 m the La Milpa Polity (LMP) in
northwest Belize spanning the Late Preclassic (400 BCE – CE 250) to the
Terminal Classic (CE 810 – 900), thus representing some of the
earliest, preserved inhabitants of the region. Burial variation exists
throughout regional occupation history, but it is unclear whether these
differences result from migration, kinship, or something else..
m the La Milpa Polity (LMP) in
northwest Belize spanning the Late Preclassic (400 BCE – CE 250) to the
Terminal Classic (CE 810 – 900), thus representing some of the
earliest, preserved inhabitants of the region. Burial variation exists
throughout regional occupation history, but it is unclear whether these
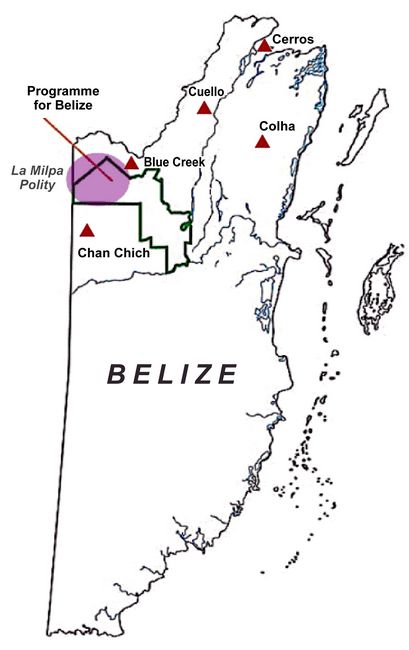
differences result from migration, kinship, or something else.. Fig.1: Relation of the Rio Bravo Conservation and Management Area (RBCMA) to the country of Belize. The purple circle indicates the suggested boundaries of the La Milpa Polity. (Image courtesy of the Programme for Belize Archaeological Project.)
Previous Work at the Site:
This research builds upon archaeological data collected from the LMP over the course of 25 field seasons. The LMP is situated within the Rio Bravo Conservation and Management Area (RBCMA), a private reserve in northwestern Belize that encompasses 260,000 acres of land, owned and operated by the non-governmental organization The Programme for Belize (PfB) (fig.1) (Valdez and Cortes-Rincon 2012). Occupation in the RBCMA spans from the Paleoindian Period (15,000 – 8,000 BCE) through the Historical Period (Valdez and Cortes-Rincon 2012; Houk 1996). However, this research focuses on migration within the region spanning the Late Preclassic (400 BCE – 250 CE) to the Terminal Classic (CE 810 – 900). Human migration has been at the forefront of explaining population ebbs and flows throughout regional occupation history, with people moving into the LMP during the Late Preclassic, depopulating in the Early Classic (CE 250 – 550), repopulating in the Late Classic (CE 550 – 810), and then abandoning the region in the Terminal Classic. However, because no research had yet been completed to test these hypotheses, this study is the first to investigate the role migration played in these population changes through time.
Approximately 150 individuals have been recovered from the project, encompassing 12 ancient Maya communities. Drake (2016) and Geller (2004) provided robust databases of information from RBCMA burials, analyzing patterns regarding site type, time period, body position, age-at-death, and sex of the decedents. The burials investigated by Drake (2016) and Geller (2004) generally possessed similar characteristics, with single-individual interments in which the decedent was often placed in a flexed position on the left side. Bodies were most commonly oriented north-south, with the head to the south however, variation exists in body positioning and orientation. These variations have been used to support hypotheses of the presence of immigrant populations. Of the patterns observed by Drake (2016) and Geller (2004), burial treatments of individuals from the RBCMA do not appear to differ greatly regarding sex of the decedents.
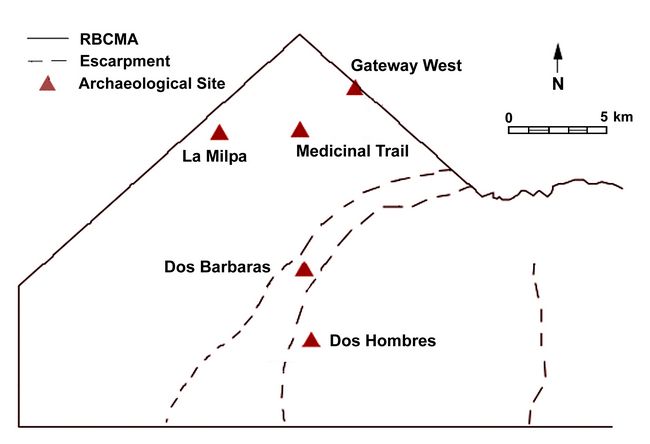 We selected 49 individuals from two
rural, one mid-sized, and two urban communities within the La Milpa
polity for this research (see Tables 1 and 2; see also fig.2). Each
community is represented by one to three households, with a minimum of
seven individuals per community and three individuals per
household. These individuals were exported to the University of
Texas at Austin for isotopic and genetic analyses.
We selected 49 individuals from two
rural, one mid-sized, and two urban communities within the La Milpa
polity for this research (see Tables 1 and 2; see also fig.2). Each
community is represented by one to three households, with a minimum of
seven individuals per community and three individuals per
household. These individuals were exported to the University of
Texas at Austin for isotopic and genetic analyses. Fig 2: Relation of the sites included in this study to one another. Adapted from Houk 1996. Image courtesy of the Programme for Belize Archaeological Project.
2018 Funded Research Project [RFF-2018-51]
Goals:
1) Identify non-local individuals from a given sample population.
2) Identify any burials patterns of non-local and local individuals and assess whether differences exist between the groups.
3) Investigate how individuals are related to one another within a single household, a household group, and a community.
Sample Selection:
M1 molars were preferentially selected (n=35); however, when not obtainable, we selected M2 (n=5) or M3 (n=9) molars, based on availability. Molars were only selected from burials in which 1) teeth were present and 2) more than two molars were available. In the instance that less than two molars were recovered, we declined to include the individual in the population sample. All teeth have been photographed and 3-D printed to preserve information for future research.
Molars are the ideal sample source: 1) preservation in the neotropics is typically poor, and human teeth often represent the most well-preserved aspect of the available specimens from the RBCMA. 2) Molars form during a relatively similar time interval in the lives of individuals; hence, the measured oxygen isotope ratios stem from similar ages across the mortuary population, regardless of age at time of death (Hodell et al. 2004; Wright 2005; Bentley 2006; Somerville 2010; Freiwald 2011; Sutinen 2014). 3) Unlike bone, human tooth enamel is highly resistant to diagenetic influences in isotopic values (Wright 2005; Wright et al. 2010). 4) Teeth have been shown to preserve aDNA better that most bone samples (Adler et al. 2011).
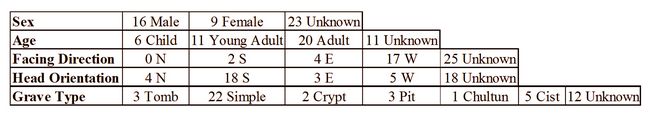 Table 1: Osteological data for individuals included in this research.
Table 1: Osteological data for individuals included in this research. Methods:
This research investigates regional and temporal processes of migration and relatedness among the ancient Maya from the PfBAP area. Forty-nine individuals were selected from the PfBAP burial population from the sites of Dos Hombres, La Milpa, Dos Barbaras, the Gateway site, and Medicinal Trail. Molars from these selected individuals were exported to The University of Texas at Austin for isotopic analyses and ancient DNA extractions and analyses.
Oxygen Isotope Background:
Oxygen isotopes are representative of local meteoric rainwater and vary depending upon the distance travelled from the ocean, cooling history of the air, altitude, humidity, and precipitation patterns (White et al. 1998, 2000, 2002, 2004a, 2007; Evans et al. 2006a; Eckardt et al. 2009; Price et al. 2010; Wright et al. 2010). Moreover, oxygen isotopes are susceptible to biological and environmental processes (Webb et al. 2013; Symonds et al. 2014). It is possible that changing environmental factors, such as hurricanes, heavy flooding, and evaporation during the dry season affected the δ18O values of water collected and stored in ancient Maya aguadas and depressions. Evaporation during dry seasons can increase δ18O values in standing water (like aguadas) and waxy leaves.
Due to these inconsistencies, baseline values for oxygen isotopes are statistically calculated (Lightfoot et al. 2016) and not reliant upon directly measured proxies. Faunal remains are not a reliable proxy, because animals often obtain their water much differently than humans (Dupras and Schwarcz 2001). Furthermore, values are not representative across species. Fractionation occurs during species-specific bone mineralization and evaporation effects on oxygen from plants consumed as water resources by animals result in different δ18O values (Dupras and Schwarcz 2001; Wright et al. 2010).
Additionally, meteoric water is not the only contributor to oxygen values in humans. Breastfeeding has proven to have an influence on the δ18O values in infants and consequently teeth that form during infancy, like incisors and canines. Breastmilk forms from body water, not meteoric waters, and thus has a heavier isotopic value (White et al. 2002, 2004a, 2007; Eckardt et al. 2009). Scholars suggest the weaning process occurred between the ages of 2 and 4 years in the Maya region (White et al. 2002, 2004a, 2007), so samples can be selected to avoid isotopic influence resulting from breastfeeding.
Furthermore, humans’ δ18O values are depleted through sweat, urine, feces, and carbon dioxide (White et al. 2000, 2002, 2004a, 2004b, 2007; Dupras and Schwarcz 2001; Price et al. 2012). As a result, statistical analyses—median absolute deviation from the median (MDM) and interquartile ranges (IQ)—must be completed on burials from each site and from each time period to try to obtain representative local baseline values. Research suggests variation within a given population is typically less than 2‰ (Price et al. 2012).
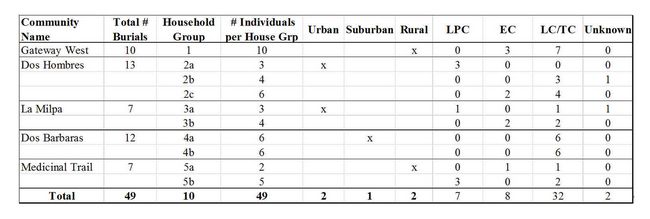 Table
2: Listing of sites and the number of respective burials included in
this research. LPC = Late Preclassic (400 BCE – CE 250); EC = Early
Classic (CE 250 – 550); LC/TC = Late Classic to Terminal Classic (CE
550 - 900). While these are generally classified as two separate time
periods, ceramic and burial data from the RBCMA is not always robust
enough to distinguish between the two.
Table
2: Listing of sites and the number of respective burials included in
this research. LPC = Late Preclassic (400 BCE – CE 250); EC = Early
Classic (CE 250 – 550); LC/TC = Late Classic to Terminal Classic (CE
550 - 900). While these are generally classified as two separate time
periods, ceramic and burial data from the RBCMA is not always robust
enough to distinguish between the two.Oxygen Isotope Sample Preparation:
Prior to wet chemistry, tooth surfaces were abraded approximately 1mm using a tungsten carbide bur. Samples were ultrasonically cleaned in 18 megOhm (D2) water for 1 hour and dried at 50˚C. The process was repeated until the water remained clear after ultrasonic cleansing. Approximately 20mg of enamel was drilled from each tooth for oxygen isotope analysis.
Sample preparation was completed following protocols outlined by Stephan (2000), Meier-Augenstein (2010), and Grimes and Pellegrini (2013). All samples were prepared in The Center for Stable Isotope Mass Spectrometry at The University of Texas at Austin. Approximately 20mg of dental enamel powder was soaked in 2.5% NaOCl for 24 hours, centrifuged, and rinsed with DI water until chloride free. Samples were soaked in 0.125M NaOH for 48 hours, centrifuged, and rinsed with DI water until pH neutral. Samples were then dissolved in 2M HF for 24 hours, reprecipitated as silver orthophosphate (Ag3PO4) with a silver ammine buffer, filtered with 0.45µm filters, and dried at 50˚C for 12 hours. Approximately 0.8mg of each sample was placed into 3.5x5mm silver capsules which were then tightly folded and stored in a vacuum oven at 70˚C for up to one week before analysis.
Results and Discussion:
Oxygen Isotopes:
The bioapatite isotopic data may indicate 38% of the sample population represent immigrants in the region. However, dependent upon the local variation in waters, our immigrant population may drop to 18%. Instead, we may have identified intra-site migration based on water dependencies of the past inhabitants, such that we may be able to recognize sites that were able to solely sustain on river water sources vs sites that depended much more frequently on water catchments and aguadas. Isotopic data do not support previous hypotheses (Welsh 1988) that the ancient Maya had regional burial practices, such that immigrants could be identified by burials patterns. Body orientation and body positioning do not correlate to immigrant or local, site type (e.g. rural or urban), or time period. Instead, we see variation in burial practices across all site types and times regardless of place of birth.
For oxygen, the determination of local has proven to be complex. Typically, non-local individuals are identified through statistical means, and while the literature typically suggests populations do not generally carry more than a 2‰ (per mil) difference, our data may suggest a much larger variability within the local range for oxygen isotope ratios. To date, we have analyzed 50 groundwater samples from the PfBAP research area. An additional 50 samples—collected from local streams, rivers, and precipitation this past summer—are currently undergoing analysis to better define the local oxygen variability within the region, elucidate seasonal variations in isotopic signatures, and illuminate the impact evapotranspiration and evaporation have on various water reservoirs the ancient Maya would have depended on for drinking water. We have positively identified non-local individuals at each site; however, the total percentage of non-local individuals may shift depending upon the returned isotopic data of local waters. We do not suggest local waters act as a proxy for ancient isotope ratios; rather, we are interested in identifying any fluctuation that may help explain the wide range in oxygen delta values from the sampled individuals.
aDNA Analysis:
In addition to identifying individual immigrants via isotopic analyses, we have successfully completed genetic analyses on the burial sample population from PfBAP. Genomics research will allow us to examine kinship, kinship practices, ancestral veneration, long term population genetic shifts, adaptations, and relatability between populations. The existing genomics research for the ancient Maya is severely limited. We are aware of only three publications which discuss original research for Ancient Maya genetics (González-Oliver et al. 2001; Iglesias et al. 2001; Merriwether et al. 1997); and only a slightly larger number discuss modern Maya genetics (e.g. Gómez-Casado et al. 2003; Herrera et al. 2007; Ibarra-Rivera et al. 2008; Martínez-González et al. 2012; Söchtig et al. 2015).
Of the 15 samples analyzed in this preliminary dataset for ancient DNA, 9 yielded endogenous Native American DNA sequences from haplogroups A2 and C1. We obtained funding for the remaining 34 individuals and are finishing up library preparations to continue our investigations regarding kinship. Furthermore, we will select 10 individuals for shotgun sequencing in an attempt to capture nuclear genome data. We are extremely excited about the potential for the aDNA research as the data we have uncovered will more than double the existing ancient Maya genetic data available and may offer some of the first mitogenome data for the ancient Maya, as earlier studies focused solely on HVR-1 sequences (González-Oliver et al. 2001; Iglesias et al. 2001; Merriwether et al. 1997).
Conclusion:
Migration in the LMP does not appear to have varied over time. Immigrants have been identified at every site included in this study and statistically represent similar percentages, indicating immigration was constant through time. We suggest that immigration does not explain regional population ebbs and flows, as originally suggested, nor do burial patterns appear to be indicative of immigrant status. To investigate these anomalies, we are continuing to pursue genetic analyses on the sample population in an attempt to parcel out social memory, placemaking, and ancestral veneration practices of the living which may shed light on differences in burial patterning of the dead.
References:
Assmann, Jan and John Czaplicka
1995 Collective Memory and Cultural Identity. New German Critique 65: 125-133.
Chesson, Meredith S.
2001 Social Memory, Identity, and Death: An Introduction. Archaeological Papers of the American Anthropological Association 10(1): 1-10.
Connerton, Paul
1989 How Societies Remember. Cambridge University Press, Cambridge
Drake, Stacy Marie
2016 Regional Perspective of Ancient Maya Burial Patterns in Northwest Belize, Central America. Ph.D. dissertation, Department of Anthropology, The University of Texas at Austin.
Geller, Pamela L.
2004 Transforming Bodies, Transforming Identities: A Consideration of Pre-Columbian Maya Corporeal Beliefs and Practices. Ph.D. dissertation, Department of Anthropology, University of Pennsylvania.
Gillespie, Susan D.
2010 Maya Memory Work. Ancient Mesoamerica 21: 401-414.
Gómez-Casado, E., J. Martínez-Laso, J. Moscoso, J. Zamora, M. Martin-Villa, M. Perez-Blas, M. Lopez-Santalla, P. Lucas Gramajo, C. Silvera, E. Lowy, A. Arnaiz-Villena
2003 Origin of Mayans According to HLA Genes and the Uniqueness of Amerindians.Tissue Antigens 61: 425-436.
González-Oliver, Angélica, Lourdes Márquez-Morfín, José C. Jiménez, and Alfonso Torre-Blanco
2001 Founding Amerindian Mitochondrial DNA Lineages in Ancient Maya from Xcaret, Quintana Roo. American Journal of Physical Anthropology 116: 230-235.
Grimes, Vaughan and Maura Pellegrini
2013 A Comparison of Pretreatment Methods for the Analysis of Phosphate Oxygen Isotope Ratios in Bioapatite. Rapid Communications in Mass Spectrometry 27: 375-390.
Halbwachs, Maurice
1980 The Collective Memory, translated by Francis J. Ditter, Jr. and Vida Yazdi Ditter. Harper and Row, New York.
Herrera, R.J., D.P. Rojas, M.C. Terreros
2007 Polymorphic Alu Insertions among Mayan Populations. Journal of Human Genetics52: 129-142.
Houk, Brett Alan
1996 The Archaeology of Site Planning: an Example from the Maya Site of Dos Hombres, Belize. Ph.D. dissertation, Department of Anthropology, The University of Texas at Austin.
Ibarra-Rivera, Lisa, Sheyla Mirabal, Manuela M. Regueiro, and Rene J. Herrera
2008 Delineating Genetic Relationships among the Maya. American Journal of Physical Anthropology 135: 329-347.
Iglesias, María Josefa, Andrés Ciudad, Eduardo Arroyo, Jesús Adánez, Sara Álvarez
2001 Aplicaciones de la Antropología Molecular a la Arqueología Maya: El Caso de Tikal. In XIV Simposio de Investigaciones Arqueológicas en Guatelama, 2000, Edited by J.P. Laporte, A.C. Suasnávar, and B. Arroyo, pp. 919-938. Museo Nacional de Arqueología y Etnología, Guatemala.
Martínez-González, Luis J., María Saiz, María J. Álvarez-Cubero, Antonio Gómez-Martín, Juan C. Álvarez, Cristina Martínez-Labarga, José A. Lorente
2012 Distribution of Y Chromosomal STRs Loci in Mayan and Mestizo Populations from Guatemala. Forensic Science International: Genetics 6: 136-142.
Meier-Augenstein, Wolfram
2010 Stable Isotope Forensics: An Introduction to the Forensic Application of Stable Isotopes. Wiley-Blackwell: Chichester, West Sussex.
Merriwether, D. Andrew, David M. Reed, and Robert E. Ferrell
1997 Ancient and Contemporary Mitochondrial DNA Variation in the Maya. In Bones of the Maya: Studies of Ancient Skeletons, Edited by, Stephen L. Whittington and David M. Reed, pp. 208-217. Smithsonian Institution Press: Washington, D.C.
Söchtig, Jens, Vanesa Ãlvarez-Iglesias, Ana Mosquera-Miguel, Miguel Gelabert-Besada, Alberto Gómez-Carballa, and Antonio Salas
2015 Genomic Insights on the Ethno-History of the Maya and the ‘Ladinos’ from Guatemala. BMC Genomics 16(1): 131.
Stephan, E.
2000 Oxygen Isotope Analysis of Animal Bone Phosphate: Method Refinement, Influence of Consolidants, and Reconstruction of Palaeotemperatures for Holocene Sites. Journal of Archaeological Science 27: 523-535.
Valdez, Fred and Marisol Cortes-Rincon
2012 Background and Introduction to the 2011 Season of the Programme for Belize Archaeological Project. In Research Reports from the Programme for Belize Archaeological Project, Volume Six, edited by Marisol Cortes-Rincon and Fred Valdez, Jr., pp1-4. Occasional Papers, Number 14. Mesoamerican Archaeological Research Laboratory, The University of Texas at Austin.
Welsh, W.B.M.
1988 An Analysis of Classic Lowland Maya Burials. BAR International: Oxford, England.
Recent Foundation grants: general Archaeology Grants Program w/map
Copyright © 2019 Rust Family Foundation