Rust Family Foundation: Archaeology Grants Program
Contextualizing
Middle Stone Age Behavioral and Cognitive Development: Insights from
Herbivore Isotope Ecology at Sibudu, KwaZulu-Natal, South Africa
Principal Investigator: Joshua R. Robinson
Instructor, Department of Anthropology, University of South Carolina
Importance of the site and project:
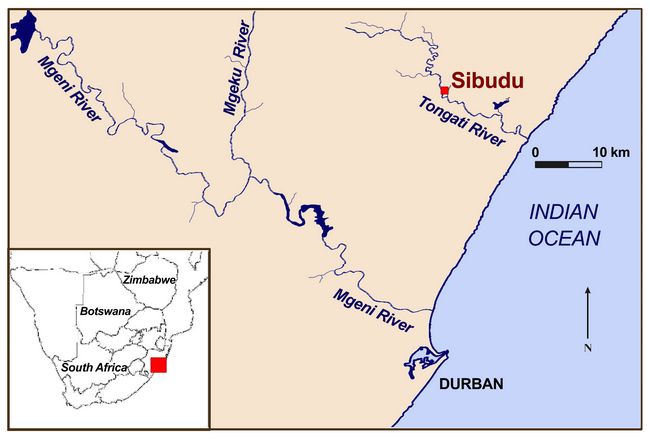
Sibudu is located on a forested cliff 20 meters above the uThongathi River approximately 40 km north of Durban and 15 km inland from the Indian Ocean in KwaZulu-Natal, South Africa (Fig.1). Of the known South African Middle Stone Age (MSA) sites, Sibudu is unique in terms of having a detailed, well-dated, and extensive MSA sequence spanning from the pre-Still Bay (before ~77,000 years before present (BP)) to the final MSA (~38,000 years BP).
 While
hiatuses do exist between these periods, the fact that several MSA
industries are present and were systematically excavated is rare.
While
hiatuses do exist between these periods, the fact that several MSA
industries are present and were systematically excavated is rare. Fig.1: Location of Sibudu near Durban in KwaZulu-Natal Province, South Africa. Inset shows southern Africa.
Excellent preservation of seeds, bone, and other organic materials—also uncommon at MSA sites—has allowed for multi-faceted environmental reconstructions for the MSA techno-complexes represented at the site. As such, Sibudu provides an opportunity to explore the extent to which the environment acted as a selective pressure for the behavioral variability and cognitive development evidenced during the MSA. This study extends the environmental reconstructions and investigations of cultural change at Sibudu using stable isotope data. High-resolution carbon and oxygen isotope data collected from faunal tooth enamel are used to address two intertwined areas of interest. First, this analysis will help quantify, and more finely characterize, the nature of environmental and climatic transitions between the MSA industries at Sibudu. Second, isotope data will be used to investigate whether such shifts correlate with cultural changes.
Previous Work at the Site:
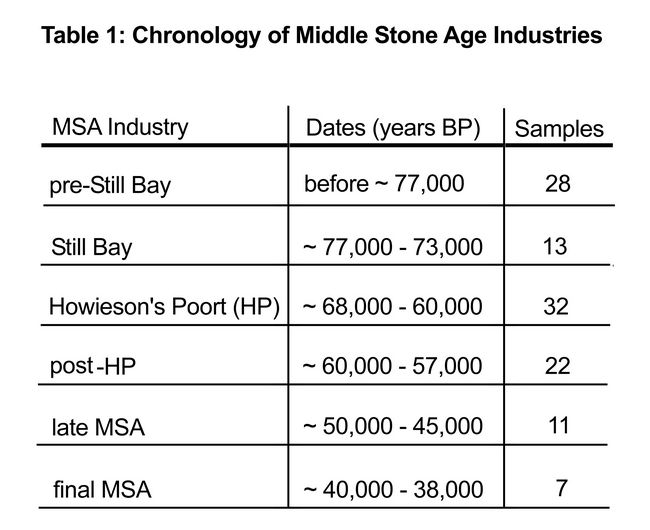
Excavation and post-excavation analyses have led to the production of an abundant literature related to the archaeological sequence, dating, lithic technology, faunal assemblage, botanical remains, pigments, hearths, and charcoal. The site preserves six MSA industries: the pre-Still Bay (before ~77,000 years BP), the Still Bay (~77,000 – 73,000 years BP), the Howieson’s Poort (HP; ~68,000 – 60,000 years BP), the post-HP (~60,000 – 57,000 years BP), the late MSA (~50,000 – 45,000 years BP), and the final MSA (~40,000 – 38,000 years BP (table 1).
 Associated
faunal and botanical remains provide some indication of habitat and
environmental change through the sequence. The HP fauna is
predominantly comprised of species that preferentially inhabit closed
environments (particularly forests), while the fauna from the
post-HP suggests a more open, savanna-like environment. The
decline of
fauna preferring closed habitats, dominated by blue duiker (Philantomba
monticola), through the HP sequence and into the post-HP is
statistically significant.
Associated
faunal and botanical remains provide some indication of habitat and
environmental change through the sequence. The HP fauna is
predominantly comprised of species that preferentially inhabit closed
environments (particularly forests), while the fauna from the
post-HP suggests a more open, savanna-like environment. The
decline of
fauna preferring closed habitats, dominated by blue duiker (Philantomba
monticola), through the HP sequence and into the post-HP is
statistically significant. Charcoal data also reveal a turnover from closed, forested environments during the HP to more open, dry conditions in the post-HP. Preserved seeds in the HP are dominated by sedges (Cyperaceae), implying the presence of standing water likely associated with the uThongathi River. Combined with isotopic analysis of Yellowwood (Podocarpus) charcoal, the botanical remains suggest that the HP at Sibudu may have been characterized by warm and wet conditions. The late MSA and final MSA units are represented by more species which prefer open habitats, including zebra (Equus quagga), continuing the trend of open habitats. The stable isotope study investigated habitat and environmental changes throughout the Sibudu sequence in order to further contextualize the appearance of the HP out of the Still Bay and its disappearance into the post-HP.
2017 Funded Research Project (RFF-2017-31):
Goals:
The aims of the study were as follows:
1. First and foremost, this study was intended to provide the first detailed on-site habitat profile of the South African MSA, particularly for the pre-50,000 years BP period.
2. Characterize whether and, if so, how, site-level environment and habitat changes between the Middle Stone Age techno-complexes at Sibudu.
3. Use the isotope data to investigate if and how environmental and habitat changes influenced cultural and technological development, particularly between the Still Bay, HP, and post-HP industries.
Methodology:
A total of 113 fossil herbivore teeth from the Sibudu archaeological collections housed at the Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa were sampled for stable carbon and oxygen isotope analysis (fig.2). Sampling focused on molars, preferably third molars, to limit the potential for interference in the dietary signal from weaning. All taxa for which securely identified molars were available were sampled. Only a single tooth from partial or complete tooth rows was sa
 mpled to avoid sampling
multiple teeth from the same individual. Sampling of teeth and export
of powdered enamel samples were conducted with a South African Heritage
Resources Agency (SAHRA) permit.
mpled to avoid sampling
multiple teeth from the same individual. Sampling of teeth and export
of powdered enamel samples were conducted with a South African Heritage
Resources Agency (SAHRA) permit.Fig.2: Sampling of herbivore tooth enamel from Sibudu for carbon and oxygen stable isotope analysis. University of the Witwatersrand, Johannesburg, South Africa.
The carbonate fraction of tooth enamel is minimally affected by diagenesis and has been shown to preserve in vivo biological signals which can be used as proxy data for diet and drinking behavior, and, in turn, paleoenvironments. Stable carbon isotope analysis allows for differentiating between C3 (trees, shrubs, and high-altitude grasses above ~ 2000 m) and C4 (most low elevation tropical grasses in Africa) photosynthetic sources of herbivore diet. The generally non-overlapping carbon (d13C) isotopic ranges for the two photosynthetic pathways in Africa are ideal for differentiating between browsers, grazers, and mixed-feeders, based on broad dietary strategies. Stable oxygen isotope composition has been used as a proxy for aridity with the assumption that during arid periods the lighter oxygen isotope, 16O, will be preferentially lost in evaporation relative to the heavier, 18O, isotope. The result of this scenario is heavier, or 18O enriched, signals recorded in faunal tooth enamel.
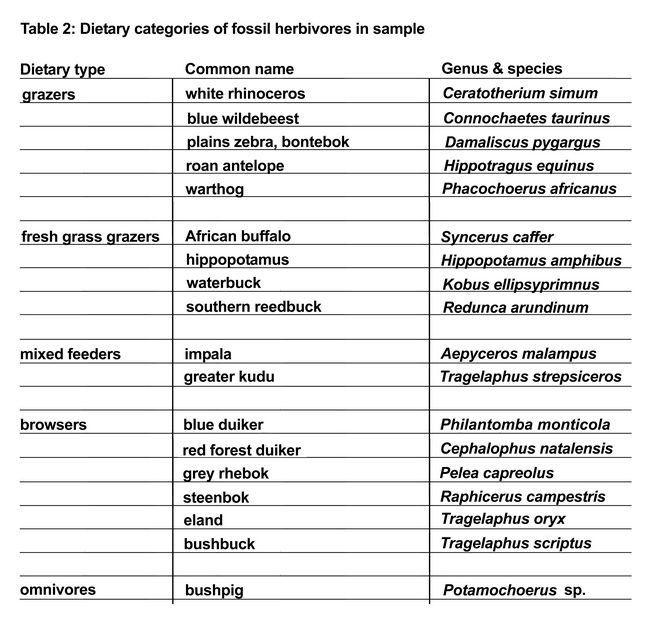
In addition to descriptive and statistical analyses of individual taxa, species are also grouped into dietary categories for further statistical analyses with larger sample sizes. Dietary categories are defined based on observational and published isotope data and are as follows (table 2): grazers – white rhinoceros (Ceratotherium simum), blue wildebeest (Connochaetes taurinus), plains zebra, bontebok (Damaliscus pygargus), roan antelope (Hippotragus equinus), and warthog (Phacochoerus africanus); fresh grass grazers – African buffalo (Syncerus caffer), hippopotamus (Hippopotamus amphibus), waterbuck (Kobus ellipsyprimnus), and southern reedbuck (Redunca arundinum); mixed-feeders – impala (Aepyceros melampus) and greater kudu (Tragelaphus strepsiceros); browsers – blue duiker (Philantomba monticola), red forest duiker (Cephalophus natalensis), grey rhebok (Pelea capreolus), steenbok (Raphicerus campestris), eland (Tragelaphus oryx), and bushbuck (Tragelaphus scriptus); omnivores – bushpig (Potamochoerus sp.).
For each taxon within a dietary category, 95% confidence intervals (CI) for both mean d13C and oxygen (d18O) were calculated through bootstrapping with replacement over 10,000 iterations. While d13C and d18O are broadly overlapping for most taxa within each of these dietary categories, white rhinoceros has lower d13C values than other grazers, southern reedbuck has higher d18O values than other fresh grass grazers, and steenbok has higher d13C values than the other browsers. As such, these taxa are excluded from their respective dietary categories for statistical analyses.
Results of 2017 project:
African buffalo (n=22; mean: 3.4±4.3‰) with its high isotopic variability is the only species which indicates differences in d13C values through the Sibudu sequence, although a Kruskal-Wallis test across all six MSA techno-complexes fails to find this significant (p=0.078). The general trend through the sequence is towards higher d13C values over time with a Mann-Whitney U test (p=0.017) finding the pre-Still Bay (n=7; mean: 5.7±3.6‰) to have significantly lower d13C values than the late MSA (n=3; mean: 0.9±0.3‰). No significant differences are found between the Still Bay (n=3; mean: 5.0±3.9‰) and the HP (n=4; p=1.000; mean: 3.8±2.9‰), or the HP and the Post-HP (n=3; p=0.857; mean: 3.6±7.2‰). The other two most commonly sampled taxa in the Sibudu sequence are blue duiker (n=21; mean: 12.2±1.8‰) and bushpig (n=32; mean: 14.1±3.3‰). Neither Kruskal-Wallis tests (blue duiker: p=0.315; bushpig: p=0.214) nor any pair-wise Mann-Whitney U tests find significant differences in the Sibudu sequence.
Estimates of C4 consumption based on d13C values further supports a shift in African buffalo diet through the Sibudu sequence from consuming ~40-45% C4 resources in the pre-Still Bay and Still Bay periods to ~50% in the HP and post-HP to ~80% and ~90% in the late MSA and final MSA, respectively. Only the late MSA and final MSA are comparable to a modern sample (n=117) from eastern Africa which indicates ~80% C4 resource consumption. Wildebeest, which prefer grazing in more open habitats, maintain ~80% C4 resource consumption throughout the sequence, although a single sample from HP is estimated to have consumed only ~60% C4 vegetation. In the HP, the dedicated grazer roan antelope is estimated to have consumed only 55% C4 resources (n=4). Roan antelope (n=1; 100%), wildebeest (n=4; 90%), and zebra (n=5; 70%) are estimated to have consumed C4 resources more in line with modern analogs from the Post-HP through the final MSA.
Analysis of dietary categories generally supports the trends for individual species. None of the dietary categories are characterized by significant trends over time as determined by Kruskal-Wallis tests (grazers: n=20, p=0.144; fresh grass grazers: n=25, p=0.250; browsers: n=27, p=0.207). Mann-Whitney U tests indicate some differences between MSA industries. For grazers, the late MSA (n=3; mean: 1.0±1.8‰) has significantly higher d13C values (p=0.033) than the HP (n=8; mean: 4.3±5.0‰). The late MSA (n=3; mean: 0.9±0.3‰) also has significantly higher d13C values than the pre-Still Bay (n=7; mean: 5.7±3.6‰; p=0.017) and the Still Bay (n=3; mean: 5.0±3.9‰; p=0.049) for fresh grass grazers.
Similar to d13C, African buffalo is the only taxon which suggest change in d18O values through the Sibudu sequence. A Kruskal-Wallis test (p=0.018) indicates a significant trend which is toward higher d18O values over time (note that sample sizes presented in Section 3.2 are not repeated in this section). Mann-Whitney U tests reveal that lower d18O values in the pre-Still Bay (mean: 5.3±0.5‰) are driving this trend. The pre-Still Bay is characterized by significantly lower d18O values for African buffalo than the Still Bay (p=0.017; mean: 3.5±0.6‰), the HP (p=0.012; mean 4.0±0.9‰), and the late MSA (p=0.033; mean: 2.8±1.9‰). Again, Mann-Whitney U tests fail to find significant differences between the Still Bay and HP (p=0.629) or the HP and the Post-HP (p=0.857; mean: 4.2±1.7‰).
Kruskal-Wallis tests fail to identify a significant trend over time for any of the dietary categories (browsers: p=0.343), although both grazers (p=0.054) and fresh grass grazers (p=0.066) approach significance. For the fresh grass grazers, which are dominated by African buffalo, this trend appears to be driven by lower d18O values during the pre-Still Bay. For grazers, the trend is driven by higher d18O values during the late MSA (mean: 0.9±1.0‰), which Mann-Whitney U tests show are significantly higher than the HP (mean: 3.9±1.4‰; p=0.017) or the post-HP (n=9; mean: 4.3±1.9‰; p=0.036). No differences are found between the HP and post-HP (p=0.606) for grazers. There are no grazer samples from the Still Bay.
Conclusions:
The South African Middle Stone Age record, and particularly the appearance and disappearance of the Still Bay and HP techno-complexes, has led to decades of debate as to whether it is the result of climatic and environmental conditions or changes in social and demographic factors. Few studies have considered scale-appropriate, on-site testing of behavior-environment links in reference to South Africa. Two periods of habitat change are identified at Sibudu. The pre-Still Bay (> 77,000 years BP) is characterized by more closed, likely heavily forested, and mesic conditions than the rest of the sequence. Late and final Middle Stone Age industries (~ 50,000 – 38,000 years BP) at Sibudu are associated with more open and likely drier conditions than earlier.
No major environmental or habitat changes based on stable isotope data, however, are identified in the Sibudu record between the Still Bay, HP, or post-HP techno-complexes. At least at Sibudu, it appears as if demographic factors may be more at play than habitat changes in Middle Stone Age technological transitions. The Still Bay – HP – Post-HP portion of the sequence is characterized by a relatively stable closed, likely forested, and mesic environment. Environmental factors, however, do seem to be associated with the end of the Middle Stone Age at Sibudu, perhaps signaling the onset of the Later Stone Age in South Africa. Further generation of local, site-level, paleoenvironmental databases will produce a clearer picture of the roles of and relationships between climatic and demographic factors in the development of Middle Stone Age behaviors in South Africa.
The finding of local environmental conditions different from, or at least not easily deduced from, models based on regional climatic datasets exposes a critical need to develop additional high-resolution on-site habitat reconstructions. This is especially true of the pre-50,000 year BP period in South Africa which encompasses the Still Bay and HP. While the results from Sibudu suggest that social and demographic factors likely played a large role in these industries, data from other sites are needed to determine whether that is the case overall. To this end I am exploring additional opportunities to generate site-specific stable isotope datasets.
References:
Backwell, L., d’Errico, F., Wadley, L. (2008). Middle Stone Age bone tools from the Howiesons Poort layers, Sibudu Cave, South Africa. Journal of Archaeological Science. 35: 1566-1580.
Backwell, L., Bradfield, J., Carlson, K.J., Jashashvili, T., Wadley, L. d'Errico, F. (2018). The antiquity of bow-and-arrow technology: evidence from Middle Stone Age layers at Sibudu Cave. Antiquity, 92(362): 289-303.
Bentsen, S.E. (2013). Controlling the heat: an experimental approach to Middle Stone Age pyrotechnology. South African Archaeological Bulletin, 68: 137-145.
Cain, C. (2006). Human activity suggested by the taphonomy of 60 ka and 50 ka faunal remains from Sibudu Cave. South African Humanities. 18: 241-260.
Clark, J.L. (2011). The evolution of human culture during the later Pleistocene: using fauna to test models on the emergence and nature of “modern” human behaviour. Journal of Anthropological Archaeology. 30: 273-291.
Clark, J.L. (2013). Exploring the relationship between climate change and the decline of the Howieson’s Poort at Sibudu Cave (South Africa). In: Clark, J.L., Speth, J.D. (Eds.), Zooarchaeology and Modern Human Origins: Human Hunting Behavior During the Later Pleistocene. Springer, Dordrecht, pp. 9-18.
Clark, J.L. (2017). The Howieson's Poort fauna from Sibudu Cave: Documenting continuity and change within Middle Stone Age industries. Journal of Human Evolution, 107: 49-70.
Clark, J.L., Plug, I. (2008). Animal exploitation strategies during the South African Middle Stone Age: Howiesons Poort and post-Howiesons Poort fauna from Sibudu Cave. Journal of Human Evolution, 54: 886-898.
Collins, B. (2015). The taphonomy of the final Middle Stone Age from Sibudu Cave, South Africa. International Journal of Osteoarchaeology. 25: 805-815.
Collins, B. (2016). Foraging strategies during the final Middle Stone Age occupation at Sibudu Cave, South Africa. Journal of Archaeological Science: Reports, 5: 61-70.
de la Peña, P. (2015). Refining our understanding of the Howiesons Poort lithic technology: The evidence from Grey Rocky Layer in Sibudu Cave (KwaZulu-Natal, South Africa). PLoS ONE 10(12): e0143451.
de la Peña, P., Wadley, L. (2014a). New knapping methods in the Howiesons Poort at Sibudu (KwaZulu-Natal, South Africa). Quaternary International, 350: 26-42.
de la Peña, P., Wadley, L. (2014b). Quartz knapping strategies in the Howiesons Poort at Sibudu (KwaZulu-Natal, South Africa). PLoS ONE, 9: e101534.
de la Peña, P., Wadley, L. (2017). Technological variability at Sibudu Cave: The end of Howiesons Poort and reduced mobility strategies after 62,000 years ago. PLoS ONE, 12(10): 1-41. e0185845.
d'Errico, F., Backwell, L.R., Wadley, L. (2012). Identifying regional variability in Middle Stone Age bone technology: the case of Sibudu Cave. Journal of Archaeological Science, 39: 2479-2495.
Hall, G., Wadley, L. and Woodborne, S. (2014). Past environmental proxies from the Middle Stone Age at Sibudu, Kwazulu-Natal, South Africa. Journal of African Archaeology, 12(1): 7-24.
Hodgskiss, T. (2013). Ochre use in the Middle Stone Age at Sibudu, South Africa: Grinding, Rubbing, Scoring and Engraving. Journal of African Archaeology, 11: 75-95.
Jacobs, Z., Wintle, A., Duller, G., Roberts, R., Wadley, L. (2008a). New ages for the post-Howiesons Poort, late and final Middle Stone Age at Sibudu, South Africa. Journal of Archaeological Science, 35: 1790-1807.
Jacobs, Z., Roberts, R.G.., Galbraith, R.F., Deacon, H.J., Grun, R., Mackay, A., Mitchell, P.J., Vogelsang, R., Wadley, L. (2008b). Ages for the Middle Stone Age of Southern Africa: Implications for Human Behavior and Dispersal. Science, 322: 733-735.
Lombard, M. (2006). Direct evidence for the use of ochre in the hafting technology of Middle Stone Age tools from Sibudu Cave. South African Humanities. 18: 57-67.
Lombard, M. (2011). Quartz-tipped arrows older than 60 ka: further use-trace evidence from Sibudu, KwaZulu-Natal, South Africa. Journal of Archaeological Science, 38(8): 1918- 1930.
Miller, C., Sievers, C. (2012). An experimental micromorphological investigation of bedding construction in the Middle Stone Age of Sibudu, South Africa. Journal of Archaeological Science, 39: 3039-3051.
Plug, I. (2004). Resource exploitation: animal use during the Middle Stone Age at Sibudu Cave, KwaZulu-Natal. South African Journal of Science. 100: 151-158.
Plug, I. (2006). Aquatic animals and their associates from the Middle Stone Age levels at Sibudu. South African Humanities. 18(1): 289-299.
Plug, I., Clark, J.L. (2008). In the air: preliminary report on the birds from Sibudu Cave, KwaZulu-Natal, South Africa. South African Journal of Archaeological Science. Goodwin Series 10: 133-142.
Renaut, R., Bamford, M.K. (2006). Results of a preliminary palynological analysis at Sibudu Cave. Southern African Humanities, 18: .235-240.
Val, A. (2015). New data on the avifauna from the Middle Stone Age layers of Sibudu Cave, South Africa: taphonomic and paleoenvironmental implications. Quaternary International, 1-17.
Val, A., de la Peña, P., Wadley, L. (2016). Direct evidence for human exploitation of birds in the Middle Stone Age of South Africa: The example of Sibudu Cave, KwaZulu-Natal. Journal of Human Evolution, 99: 107-123.
Wadley, L. (2005). A Typological Study of the Final Middle Stone Age Stone Tools from Sibudu Cave, KwaZulu-Natal. South African Archaeological Bulletin, 60: 51-63.
Wadley, L. (2007). Announcing a Still Bay industry at Sibudu Cave, South Africa. Journal of Human Evolution, 52: 681-689.
Wadley, L. (2008). The Howieson’s Poort industry of Sibudu Cave. South African Archaeological Bulletin, Goodwin Series 10: 122-132.
Wadley, L., Jacobs, Z. (2004). Sibudu Cave, KwaZulu-Natal: Background to the excavations of Middle Stone Age and Iron Age occupations. South African Journal of Science, 100: 145-151.
Wadley, L., Jacobs, Z. (2006). Sibudu Cave: background to the excavations, stratigraphy and dating. Southern African Humanities, 18:1-26.
Wadley, L., Langejans, G. (2014). Preliminary study of scrapers around combustion features in layer SS, Sibudu, 58000 years ago. South African Archaeological Bulletin, 69: 19-33.
Wadley, L., Mohapi, M. (2008). A Segment is not a Monolith: evidence from the Howiesons Poort of Sibudu, South Africa. Journal of Archaeological Science, 35: 2594-260.
Wadley, L., Sievers, C., Bamford, M., Goldberg, P., Berna, F., Miller, C. (2011). Middle Stone Age bedding construction and settlement patterns at Sibudu, South Africa. Science 334: 1388-1391.
Wojcieszak, M., Wadley, L. (2018). Raman spectroscopy and scanning electron microscopy confirm ochre residues on 71 000-year-old bifacial tools from Sibudu, South Africa. Archaeometry. https://doi.org/10.1111/arcm.12369
Recent Foundation grants: general Archaeology Grants Program w/map
Copyright © 2018 Rust Family Foundation